

Infinite alleles model
Encyclopedia
The infinite alleles model is a mathematical model for calculating genetic mutation
s. The Japanese geneticist Motoo Kimura
and American geneticist James F. Crow
(1964) introduced the infinite allele
s model, an attempt to determine for a finite diploid population what proportion of loci would be homozygous. This was, in part, motivated by assertions by other geneticists that more than 50 percent of Drosophila
loci were heterozygous, a claim they initially doubted. In order to answer this question they assumed first, that there were a large enough number of alleles so that any mutation
would lead to a different allele (that is the probability of back mutation to the original allele would be low enough to be negligible); and second, that the mutations would result in a number of different outcomes from neutral to deleterious.
They determined that in the neutral case, the probability that an individual would be homozygous, F, was:
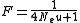
where u is the mutation rate, and Ne is the effective population size
. The effective number of alleles n maintained in a population is defined as the inverse of the homozygosity, that is
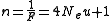
which is a lower bound for the actual number of alleles in the population.
If the effective population is large, then a large number of alleles can be maintained. However, this result only holds for the neutral case, and is not necessarily true for the case when some alleles are more or less fit
than others, for example when the fittest genotype is a heterozygote (a situation often referred to as overdominance
or heterosis
).
In the case of overdominance, because Mendel's second law
(the law of segregation) necessarily results in the production of homozygotes (which are by definition in this case, less fit), this means that population will always harbor a number of less fit individuals, which leads to a decrease in the average fitness of the population. This is sometimes referred to as genetic load
, in this case it is a special kind of load known as segregational load. Crow and Kimura showed that at equilibrium conditions, for a given strength of selection (s), that there would be an upper limit to the number of fitter alleles (polymorphisms) that a population could harbor for a particular locus. Beyond this number of alleles, the selective advantage of presence of those alleles in heterozygous genotypes would be cancelled out by continual generation of less fit homozygous genotypes.
These results became important in the formation of the neutral theory, because neutral (or nearly neutral) alleles create no such segregational load, and allow for the accumulation of a great deal of polymorphism. When Richard Lewontin
and J. Hubby published their groundbreaking results in 1966 which showed high levels of genetic variation in Drosophila via protein electrophoresis
, the theoretical results from the infinite alleles model were used by Kimura and others to support the idea that this variation would have to be neutral (or result in excess segregational load).
Mutation
In molecular biology and genetics, mutations are changes in a genomic sequence: the DNA sequence of a cell's genome or the DNA or RNA sequence of a virus. They can be defined as sudden and spontaneous changes in the cell. Mutations are caused by radiation, viruses, transposons and mutagenic...
s. The Japanese geneticist Motoo Kimura
Motoo Kimura
was a Japanese biologist best known for introducing the neutral theory of molecular evolution in 1968. He became one of the most influential theoretical population geneticists. He is remembered in genetics for his innovative use of diffusion equations to calculate the probability of fixation of...
and American geneticist James F. Crow
James F. Crow
James F. Crow is Professor Emeritus of Genetics at the University of Wisconsin–Madison.Some of his most significant peer-reviewed contributions were coauthored with Motoo Kimura. His major contribution to the field, however, is arguably his teaching...
(1964) introduced the infinite allele
Allele
An allele is one of two or more forms of a gene or a genetic locus . "Allel" is an abbreviation of allelomorph. Sometimes, different alleles can result in different observable phenotypic traits, such as different pigmentation...
s model, an attempt to determine for a finite diploid population what proportion of loci would be homozygous. This was, in part, motivated by assertions by other geneticists that more than 50 percent of Drosophila
Drosophila melanogaster
Drosophila melanogaster is a species of Diptera, or the order of flies, in the family Drosophilidae. The species is known generally as the common fruit fly or vinegar fly. Starting from Charles W...
loci were heterozygous, a claim they initially doubted. In order to answer this question they assumed first, that there were a large enough number of alleles so that any mutation
Mutation
In molecular biology and genetics, mutations are changes in a genomic sequence: the DNA sequence of a cell's genome or the DNA or RNA sequence of a virus. They can be defined as sudden and spontaneous changes in the cell. Mutations are caused by radiation, viruses, transposons and mutagenic...
would lead to a different allele (that is the probability of back mutation to the original allele would be low enough to be negligible); and second, that the mutations would result in a number of different outcomes from neutral to deleterious.
They determined that in the neutral case, the probability that an individual would be homozygous, F, was:
where u is the mutation rate, and Ne is the effective population size
Effective population size
In population genetics, the concept of effective population size Ne was introduced by the American geneticist Sewall Wright, who wrote two landmark papers on it...
. The effective number of alleles n maintained in a population is defined as the inverse of the homozygosity, that is
which is a lower bound for the actual number of alleles in the population.
If the effective population is large, then a large number of alleles can be maintained. However, this result only holds for the neutral case, and is not necessarily true for the case when some alleles are more or less fit
Fitness (biology)
Fitness is a central idea in evolutionary theory. It can be defined either with respect to a genotype or to a phenotype in a given environment...
than others, for example when the fittest genotype is a heterozygote (a situation often referred to as overdominance
Overdominance
Overdominance is a condition in genetics where the phenotype of the heterozygote lies outside of the phenotypical range of both homozygote parents. Overdominance can also be described as heterozygote advantage, wherein heterozygous individuals have a higher fitness than homozygous individuals.An...
or heterosis
Heterosis
Heterosis, or hybrid vigor, or outbreeding enhancement, is the improved or increased function of any biological quality in a hybrid offspring. The adjective derived from heterosis is heterotic....
).
In the case of overdominance, because Mendel's second law
Mendelian inheritance
Mendelian inheritance is a scientific description of how hereditary characteristics are passed from parent organisms to their offspring; it underlies much of genetics...
(the law of segregation) necessarily results in the production of homozygotes (which are by definition in this case, less fit), this means that population will always harbor a number of less fit individuals, which leads to a decrease in the average fitness of the population. This is sometimes referred to as genetic load
Genetic load
In population genetics, genetic load or genetic burden is a measure of the cost of lost alleles due to selection or mutation...
, in this case it is a special kind of load known as segregational load. Crow and Kimura showed that at equilibrium conditions, for a given strength of selection (s), that there would be an upper limit to the number of fitter alleles (polymorphisms) that a population could harbor for a particular locus. Beyond this number of alleles, the selective advantage of presence of those alleles in heterozygous genotypes would be cancelled out by continual generation of less fit homozygous genotypes.
These results became important in the formation of the neutral theory, because neutral (or nearly neutral) alleles create no such segregational load, and allow for the accumulation of a great deal of polymorphism. When Richard Lewontin
Richard Lewontin
Richard Charles "Dick" Lewontin is an American evolutionary biologist, geneticist and social commentator. A leader in developing the mathematical basis of population genetics and evolutionary theory, he pioneered the notion of using techniques from molecular biology such as gel electrophoresis to...
and J. Hubby published their groundbreaking results in 1966 which showed high levels of genetic variation in Drosophila via protein electrophoresis
Electrophoresis
Electrophoresis, also called cataphoresis, is the motion of dispersed particles relative to a fluid under the influence of a spatially uniform electric field. This electrokinetic phenomenon was observed for the first time in 1807 by Reuss , who noticed that the application of a constant electric...
, the theoretical results from the infinite alleles model were used by Kimura and others to support the idea that this variation would have to be neutral (or result in excess segregational load).