

Behavioral momentum
Encyclopedia
Behavioral momentum is a theory in quantitative analysis of behavior
and is a comparative metaphor
based on physical momentum
. It describes the general relation between resistance to change (persistence of behavior) and the rate of reinforcement
obtained in a given situation.
B.F. Skinner (1938) proposed that all behavior is based on the fundamental unit of behavior termed the discriminated operant. The discriminated operant, also known as the three-term contingency, is broken down into three necessary components: an antecedent discriminative-stimulus context, a response, and a reinforcing or punishing consequence. Responding occurs in the presence of the stimulus
because responding previously has resulted in a consequence in its presence.
& Grace, 2000, for a review). The positive reinforcement
contingency between the response and a reinforcing consequence governs response rates (i.e., a response–reinforcer relation) by shaping a particular special pattern of responding, according to the relative law of non-absolute effect (i.e., the matching law
; Herrnstein
, 1970). Conversely, the Pavlovian relation established between the context (as a discriminative stimulus) and the rate or magnitude (but not both) of reinforcement obtained in the context (i.e., a stimulus–reinforcer relation) governs the resistance of the behavior class to behavioral operations such as implementing an extinction contingency. Resistance to change is assessed as responding during conditions of environmental disruption (e.g., satiation, extinction
) relative to stable, pre-disruption behavior response rates.
Resistance to disruption has been considered a superior measure to stable response rates as an expression of the underlying strength of a response (Nevin, 1974). This is because drastic differences in response rates can occur by reinforcement
contingency shaping particular patterns of behavior (e.g., differential-reinforcement-of-high- or low-response-rate schedules) while reinforcement rates are equal. It is unclear, therefore, whether these differences in response rates necessarily indicate differences in the underlying strength of a response (see Morse, 1966, for a discussion).
According to behavioral momentum theory, the relation between response rates and resistance to change is analogous to the velocity
and mass
of a moving object, according to Newton
's second law of motion (Nevin, Mandell & Atak, 1983). Newton's second law states that the change in velocity of a moving object when an outside force is applied is directly related to that force and inversely related to the object's mass. Similarly, behavioral momentum theory states that the change in response rates under conditions of disruption (Bx) relative to baseline response rates (Bo) are directly related to the force or magnitude of disruption (f) and inversely related to the rate of reinforcement in a stimulus context (r):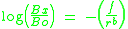 (1)
(1)
The free parameter b indicates the sensitivity of resistance to change to the rate of reinforcement in the stimulus context (i.e., the stimulus–reinforcer relation). Resistance to disruption typically is assessed when two distinctive discriminative stimulus contexts alternate and signal different schedules of reinforcement (i.e., a multiple schedule). Equation 1 can be rewritten to account for resistance to change across two stimulus contexts (Nevin, 1992; Nevin, Grace, & McLean, 2001) when a disrupter is uniformly applied across contexts (i.e., f1 = f2):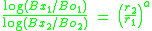 (2)
(2)
The subscripts indicate the different stimulus contexts. Thus, Equation 2 states that relative resistance to change is a power function of the relative rate of reinforcement across stimulus contexts, with the a parameter indicating sensitivity to relative reinforcement rate. Consistent with behavioral momentum theory, resistance to disruption often has been found to be greater in stimulus contexts presenting higher rates or magnitudes of reinforcement (see Nevin, 1992, for a review). Strong support for resistance to change being determined by stimulus–reinforcer relations and independent of response–reinforcer relations comes from studies that add response-independent (i.e., free) reinforcement to one stimulus context. For instance, Nevin, Tota, Torquato, and Shull (1990) had pigeons pecking lighted disks on separate variable-interval 60-s schedules of intermittent food reinforcement across two components of a multiple schedule. Additional free reinforcers were presented every 15 or 30 s on average when the disk was red, but not when the disk was green. Thus, the response–reinforcer relation was degraded when the disk was red because each reinforcer was not immediately preceded by a response. Consistent with the matching law, response rates were lower in the red context than in the green context. However, the stimulus–reinforcer relation was enhanced in the red context because the overall rate of food presentation was greater. Consistent with behavioral momentum theory, resistance to presession feeding (satiation) and discontinuing reinforcement in both contexts (extinction) was greater in the red context. Similar results have been found when reinforcers are added to a context by reinforcing an alternative response.
The findings of Nevin et al. (1990) have been extended across a number of procedures and species including goldfish (Igaki & Sakagami, 2004), rats (Harper, 1999a, 1999b; Shull, Gaynor & Grimes, 2001), pigeons (Podlesnik & Shahan, 2008), and humans (Ahearn, Clark, Gardenier, Chung & Dube, 2003; Cohen, 1996; Mace et al., 1990). The behavioral momentum framework also has been used to account for the partial-reinforcement extinction effect (Nevin & Grace, 1999), to assess the persistence of drug-maintained behavior (Jimenez-Gomez & Shahan, 2007; Shahan & Burke, 2004), to increase task compliance (e.g., Belfiore, Lee, Scheeler & Klein, 2002), and to understand the effects of social policies on global problems (Nevin, 2005).
Although behavioral momentum theory is a powerful framework for understanding how a context of reinforcement can affect the persistence of discriminated operant behavior, there are a number of findings that are inconsistent with the theory (see Nevin & Grace, 2000, and accompanying commentary). For instance, with equal reinforcement rates across stimulus contexts, resistance to change has been shown to be affected by manipulations to response–reinforcer relations, including schedules that produce different baseline response rates (e.g., Lattal, 1989; Nevin, Grace, Holland & McLean), delays to reinforcement (e.g., Bell, 1999; Grace, Schwendimann & Nevin, 1998; Podlesnik, Jimenez-Gomez, Ward & Shahan, 2006; Podlesnik & Shahan, 2008), and by providing brief stimuli that accompany reinforcement (Reed & Doughty, 2005). Also, it is unclear what factors affect relative resistance to change of responding maintained by conditioned reinforcement (Shahan & Podlesnik, 2005) or two concurrently available responses when different rates of reinforcement are arranged within the same context for those responses (e.g., Bell & Williams, 2002).
Grace and Nevin (1997) assessed both relative resistance to change in a multiple schedule and preference in a concurrent-chains procedure with pigeons pecking lighted disks for food reinforcement. When the relative rate of reinforcement was manipulated identically and simultaneously across stimulus contexts in the multiple schedule and concurrent-chains procedure, both relative resistance to change and preference was greater with richer contexts of reinforcement. When all the extant resistance to change and preference data were summarized by Grace, Bedell, and Nevin (2002), they found that those measures were related by a structural relation slope of 0.29. Therefore, relative resistance to change and preference both have been conceptualized as expressions of an underlying construct termed response strength, conditioned reinforcement value, or more generally, behavioral mass of discriminated operant behavior (see Nevin & Grace, 2000).
Quantitative Analysis of Behavior
Quantitative analysis of behavior is the quantitative form of the experimental analysis of behavior. This has become the dominant scientific approach to behavior analysis. It represents behavioral research using quantitative models of behavior. The parameters in the models hopefully have...
and is a comparative metaphor
Metaphor
A metaphor is a literary figure of speech that uses an image, story or tangible thing to represent a less tangible thing or some intangible quality or idea; e.g., "Her eyes were glistening jewels." Metaphor may also be used for any rhetorical figures of speech that achieve their effects via...
based on physical momentum
Momentum
In classical mechanics, linear momentum or translational momentum is the product of the mass and velocity of an object...
. It describes the general relation between resistance to change (persistence of behavior) and the rate of reinforcement
Rate of reinforcement
In behaviorism, rate of reinforcement is number of reinforcements per time, usually per minute. Symbol of this rate is usually Rf. Its first major exponent was B.F. Skinner . It is used in the Matching Law....
obtained in a given situation.
B.F. Skinner (1938) proposed that all behavior is based on the fundamental unit of behavior termed the discriminated operant. The discriminated operant, also known as the three-term contingency, is broken down into three necessary components: an antecedent discriminative-stimulus context, a response, and a reinforcing or punishing consequence. Responding occurs in the presence of the stimulus
Stimulus (physiology)
In physiology, a stimulus is a detectable change in the internal or external environment. The ability of an organism or organ to respond to external stimuli is called sensitivity....
because responding previously has resulted in a consequence in its presence.
Resistance to change
According to behavioral momentum theory, there are two separable aspects of the discriminated operant that independently govern the rate at which a response occurs and the persistence of that response in the face of operant disruption such as punishment, extinction, differential reinforcement of alternative behaviors, etc. (see NevinJohn Anthony Nevin
John Anthony Nevin is Professor Emeritus of Psychology, University of New Hampshire. He was born July 5, 1933, in New York City. In 1954, he obtained a B.E. in Mechanical Engineering from Yale University, in 1961 an M.A. at Columbia University, then in 1963 a Ph.D. in Psychology. William N....
& Grace, 2000, for a review). The positive reinforcement
Reinforcement
Reinforcement is a term in operant conditioning and behavior analysis for the process of increasing the rate or probability of a behavior in the form of a "response" by the delivery or emergence of a stimulus Reinforcement is a term in operant conditioning and behavior analysis for the process of...
contingency between the response and a reinforcing consequence governs response rates (i.e., a response–reinforcer relation) by shaping a particular special pattern of responding, according to the relative law of non-absolute effect (i.e., the matching law
Matching law
In operant conditioning, the matching law is a quantitative relationship that holds between the relative rates of response and the relative rates of reinforcement in concurrent schedules of reinforcement...
; Herrnstein
Richard Herrnstein
Richard J. Herrnstein was an American researcher in animal learning in the Skinnerian tradition. He was one of the founders of quantitative analysis of behavior....
, 1970). Conversely, the Pavlovian relation established between the context (as a discriminative stimulus) and the rate or magnitude (but not both) of reinforcement obtained in the context (i.e., a stimulus–reinforcer relation) governs the resistance of the behavior class to behavioral operations such as implementing an extinction contingency. Resistance to change is assessed as responding during conditions of environmental disruption (e.g., satiation, extinction
Extinction
In biology and ecology, extinction is the end of an organism or of a group of organisms , normally a species. The moment of extinction is generally considered to be the death of the last individual of the species, although the capacity to breed and recover may have been lost before this point...
) relative to stable, pre-disruption behavior response rates.
Resistance to disruption has been considered a superior measure to stable response rates as an expression of the underlying strength of a response (Nevin, 1974). This is because drastic differences in response rates can occur by reinforcement
Reinforcement
Reinforcement is a term in operant conditioning and behavior analysis for the process of increasing the rate or probability of a behavior in the form of a "response" by the delivery or emergence of a stimulus Reinforcement is a term in operant conditioning and behavior analysis for the process of...
contingency shaping particular patterns of behavior (e.g., differential-reinforcement-of-high- or low-response-rate schedules) while reinforcement rates are equal. It is unclear, therefore, whether these differences in response rates necessarily indicate differences in the underlying strength of a response (see Morse, 1966, for a discussion).
According to behavioral momentum theory, the relation between response rates and resistance to change is analogous to the velocity
Velocity
In physics, velocity is speed in a given direction. Speed describes only how fast an object is moving, whereas velocity gives both the speed and direction of the object's motion. To have a constant velocity, an object must have a constant speed and motion in a constant direction. Constant ...
and mass
Mass
Mass can be defined as a quantitive measure of the resistance an object has to change in its velocity.In physics, mass commonly refers to any of the following three properties of matter, which have been shown experimentally to be equivalent:...
of a moving object, according to Newton
Isaac Newton
Sir Isaac Newton PRS was an English physicist, mathematician, astronomer, natural philosopher, alchemist, and theologian, who has been "considered by many to be the greatest and most influential scientist who ever lived."...
's second law of motion (Nevin, Mandell & Atak, 1983). Newton's second law states that the change in velocity of a moving object when an outside force is applied is directly related to that force and inversely related to the object's mass. Similarly, behavioral momentum theory states that the change in response rates under conditions of disruption (Bx) relative to baseline response rates (Bo) are directly related to the force or magnitude of disruption (f) and inversely related to the rate of reinforcement in a stimulus context (r):
(1)The free parameter b indicates the sensitivity of resistance to change to the rate of reinforcement in the stimulus context (i.e., the stimulus–reinforcer relation). Resistance to disruption typically is assessed when two distinctive discriminative stimulus contexts alternate and signal different schedules of reinforcement (i.e., a multiple schedule). Equation 1 can be rewritten to account for resistance to change across two stimulus contexts (Nevin, 1992; Nevin, Grace, & McLean, 2001) when a disrupter is uniformly applied across contexts (i.e., f1 = f2):
(2)The subscripts indicate the different stimulus contexts. Thus, Equation 2 states that relative resistance to change is a power function of the relative rate of reinforcement across stimulus contexts, with the a parameter indicating sensitivity to relative reinforcement rate. Consistent with behavioral momentum theory, resistance to disruption often has been found to be greater in stimulus contexts presenting higher rates or magnitudes of reinforcement (see Nevin, 1992, for a review). Strong support for resistance to change being determined by stimulus–reinforcer relations and independent of response–reinforcer relations comes from studies that add response-independent (i.e., free) reinforcement to one stimulus context. For instance, Nevin, Tota, Torquato, and Shull (1990) had pigeons pecking lighted disks on separate variable-interval 60-s schedules of intermittent food reinforcement across two components of a multiple schedule. Additional free reinforcers were presented every 15 or 30 s on average when the disk was red, but not when the disk was green. Thus, the response–reinforcer relation was degraded when the disk was red because each reinforcer was not immediately preceded by a response. Consistent with the matching law, response rates were lower in the red context than in the green context. However, the stimulus–reinforcer relation was enhanced in the red context because the overall rate of food presentation was greater. Consistent with behavioral momentum theory, resistance to presession feeding (satiation) and discontinuing reinforcement in both contexts (extinction) was greater in the red context. Similar results have been found when reinforcers are added to a context by reinforcing an alternative response.
The findings of Nevin et al. (1990) have been extended across a number of procedures and species including goldfish (Igaki & Sakagami, 2004), rats (Harper, 1999a, 1999b; Shull, Gaynor & Grimes, 2001), pigeons (Podlesnik & Shahan, 2008), and humans (Ahearn, Clark, Gardenier, Chung & Dube, 2003; Cohen, 1996; Mace et al., 1990). The behavioral momentum framework also has been used to account for the partial-reinforcement extinction effect (Nevin & Grace, 1999), to assess the persistence of drug-maintained behavior (Jimenez-Gomez & Shahan, 2007; Shahan & Burke, 2004), to increase task compliance (e.g., Belfiore, Lee, Scheeler & Klein, 2002), and to understand the effects of social policies on global problems (Nevin, 2005).
Although behavioral momentum theory is a powerful framework for understanding how a context of reinforcement can affect the persistence of discriminated operant behavior, there are a number of findings that are inconsistent with the theory (see Nevin & Grace, 2000, and accompanying commentary). For instance, with equal reinforcement rates across stimulus contexts, resistance to change has been shown to be affected by manipulations to response–reinforcer relations, including schedules that produce different baseline response rates (e.g., Lattal, 1989; Nevin, Grace, Holland & McLean), delays to reinforcement (e.g., Bell, 1999; Grace, Schwendimann & Nevin, 1998; Podlesnik, Jimenez-Gomez, Ward & Shahan, 2006; Podlesnik & Shahan, 2008), and by providing brief stimuli that accompany reinforcement (Reed & Doughty, 2005). Also, it is unclear what factors affect relative resistance to change of responding maintained by conditioned reinforcement (Shahan & Podlesnik, 2005) or two concurrently available responses when different rates of reinforcement are arranged within the same context for those responses (e.g., Bell & Williams, 2002).
Preference and resistance to change
As resistance to disruption across stimulus contexts is analogous to the inertial mass of a moving object, behavioral momentum theory also suggests that preference in concurrent-chains procedures for one stimulus context over another is analogous to the gravitational attraction of two bodies (see Nevin & Grace, 2000). In concurrent-chains procedures, responding on the concurrently available initial links provides access to one of two mutually exclusive stimulus contexts called terminal links. As with multiple schedules, independent schedules of reinforcement can function in each terminal-link context. The relative allocation of responding across the two initial links indicates the extent to which an organism prefers one terminal-link context over the other. Moreover, behavioral momentum theory posits that preference provides a measure of the relative conditioned-reinforcing value of the two terminal-link contexts, as described by the contextual-choice model (Grace, 1994).Grace and Nevin (1997) assessed both relative resistance to change in a multiple schedule and preference in a concurrent-chains procedure with pigeons pecking lighted disks for food reinforcement. When the relative rate of reinforcement was manipulated identically and simultaneously across stimulus contexts in the multiple schedule and concurrent-chains procedure, both relative resistance to change and preference was greater with richer contexts of reinforcement. When all the extant resistance to change and preference data were summarized by Grace, Bedell, and Nevin (2002), they found that those measures were related by a structural relation slope of 0.29. Therefore, relative resistance to change and preference both have been conceptualized as expressions of an underlying construct termed response strength, conditioned reinforcement value, or more generally, behavioral mass of discriminated operant behavior (see Nevin & Grace, 2000).