

Limiting similarity
Encyclopedia
Limiting similarity is a concept in theoretical ecology
and community ecology that proposes the existence of a maximum level of niche
overlap between two given species
that will allow continued coexistence.
This concept is a corollary of the competitive exclusion principle
, which states that, controlling for all else, two species competing for exactly the same resources cannot stably coexist. It assumes normally-distributed resource utilization curves ordered linearly along a resource axis, and as such, it is often considered to be an oversimplified model of species interactions. Moreover, it has theoretical weakness, and it is poor at generating real-world predictions or falsifiable hypotheses. Thus, the concept has fallen somewhat out of favor except in didactic settings (where it is commonly referenced), and has largely been replaced by more complex and inclusive theories.
and paramecium
. The principle maintains that two species with the same ecological niche
s cannot stably coexist. That is to say, when two species compete for identical resource access, one will be competitively superior and it will ultimately supplant the other. Over the next half century, limiting similarity slowly emerged as a natural outgrowth of this principle, aiming (but not necessarily succeeding) to be more quantitative and specific.
Noted ecologist and evolutionary biologist David Lack said retrospectively that he had already begun to mull around with the ideas of limiting similarity as early as the 1940s, but it wasn't until the end of the 1950s that the theory began to be built up and articulated. G. Evelyn Hutchinson
's famous "Homage to Santa Rosalia" was the next foundational paper in the history of the theory. Its subtitle famously asks, "Why are there so many kinds of animals?", and the address attempts to answer this question by suggesting theoretical bounds to speciation and niche overlap. For the purposes of understanding limiting similarity, the key portion of Hutchinson's address is the end where he presents the observation that a seemingly ubiquitous ratio (1.3:1) defines the upper bound of morphological character similarity between closely related species. While this so-called Hutchinson ratio and the idea of a universal limit have been overturned by later research, the address was still foundational to the theory of limiting similarity.
MacArthur
and Levins
were the first to introduce the term 'limiting similarity' in their 1967 paper. They attempted to lay out a rigorous quantitative basis for the theory using probability theory
and the Lotka-Volterra competition equations. In doing so, they provided the ultimate theoretical framework on which many subsequent studies were based.
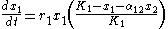
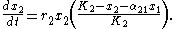
where
Macarthur and Levins examine this system applied to three populations, also visualized as resource utilization curves, depicted below. In this model, at some upper limit of competition α, between two species x1 and x3, the survival of a third species x2 between the other two is not possible. This phenomenon is termed limiting similarity. Evolutionary, if two species are more similar than some limit L, a third species will converge towards the nearer of the two competitors. If the two species are less similar than some limit L, a third species will evolve an intermediate phenotype.
May extended this theory when considering species with different carrying capacities, concluding that coexistence was unlikely if the distance between the modes of competing resource utilization curves d was less than the standard deviation of the curves w.
can be used as a close proxy for niche incongruence. One recent paleoecological
study, for example, used fossil
proxies of gastropod body size to determine levels of character displacement over 42,500 years during the Quaternary
. They found little evidence of character displacement, and they concluded that "limiting similarity, as seen in both ecological character displacement and community-wide character displacement, is a transient ecological phenomenon rather than a long-term evolutionary process". Other theoretical and empirical studies tend to find results that similarly play down the strength and role of limiting similarity in ecology and evolution. For example, Abrams (who is prolific on the subject of limiting similarity) and Rueffler find in 2009 that "there is no absolute limit to similarity; there is always some range of mortality rates of one species allowing coexistence, given a fixed mortality of the other species".
What a lot of studies examining limiting similarity find are the weaknesses in the original theory that are addressed below.
While these complications don't invalidate the concept, they render limiting similarity exceedingly difficult to test in practice and useful for little more than didacticism.
Furthermore, Hubbell and Foster point out that extinction via competition can take an extremely long time and the importance of limiting similarity in extinction may even be superseded by speciation. Also, from a theoretical standpoint, small changes in carrying capacities can allow for nearly completely overlapping resource utilization curves and in practice carrying capacity can be difficult to determine. Many studies that attempt to explore limiting similarity (including Huntley et al. 2007) resort to examining character displacement as a proxy for niche overlap, which is suspect at best. While a useful-if simple-model, limiting similarity is nearly untestable in reality.
Theoretical ecology
Theoretical ecology is the scientific discipline devoted to the study of ecological systems using theoretical methods such as simple conceptual models, mathematical models, computational simulations, and advanced data analysis...
and community ecology that proposes the existence of a maximum level of niche
Ecological niche
In ecology, a niche is a term describing the relational position of a species or population in its ecosystem to each other; e.g. a dolphin could potentially be in another ecological niche from one that travels in a different pod if the members of these pods utilize significantly different food...
overlap between two given species
Species
In biology, a species is one of the basic units of biological classification and a taxonomic rank. A species is often defined as a group of organisms capable of interbreeding and producing fertile offspring. While in many cases this definition is adequate, more precise or differing measures are...
that will allow continued coexistence.
This concept is a corollary of the competitive exclusion principle
Competitive exclusion principle
In ecology, the competitive exclusion principle, sometimes referred to as Gause's law of competitive exclusion or just Gause's law, is a proposition which states that two species competing for the same resources cannot coexist if other ecological factors are constant...
, which states that, controlling for all else, two species competing for exactly the same resources cannot stably coexist. It assumes normally-distributed resource utilization curves ordered linearly along a resource axis, and as such, it is often considered to be an oversimplified model of species interactions. Moreover, it has theoretical weakness, and it is poor at generating real-world predictions or falsifiable hypotheses. Thus, the concept has fallen somewhat out of favor except in didactic settings (where it is commonly referenced), and has largely been replaced by more complex and inclusive theories.
History
In 1932, Georgii Gause created the competitive exclusion principle based on experiments with cultures of yeastYeast
Yeasts are eukaryotic micro-organisms classified in the kingdom Fungi, with 1,500 species currently described estimated to be only 1% of all fungal species. Most reproduce asexually by mitosis, and many do so by an asymmetric division process called budding...
and paramecium
Paramecium
Paramecium is a group of unicellular ciliate protozoa, which are commonly studied as a representative of the ciliate group, and range from about 0.05 to 0.35 mm in length. Simple cilia cover the body, which allow the cell to move with a synchronous motion at speeds of approximately 12 body...
. The principle maintains that two species with the same ecological niche
Ecological niche
In ecology, a niche is a term describing the relational position of a species or population in its ecosystem to each other; e.g. a dolphin could potentially be in another ecological niche from one that travels in a different pod if the members of these pods utilize significantly different food...
s cannot stably coexist. That is to say, when two species compete for identical resource access, one will be competitively superior and it will ultimately supplant the other. Over the next half century, limiting similarity slowly emerged as a natural outgrowth of this principle, aiming (but not necessarily succeeding) to be more quantitative and specific.
Noted ecologist and evolutionary biologist David Lack said retrospectively that he had already begun to mull around with the ideas of limiting similarity as early as the 1940s, but it wasn't until the end of the 1950s that the theory began to be built up and articulated. G. Evelyn Hutchinson
G. Evelyn Hutchinson
George Evelyn Hutchinson FRS was an Anglo-American zoologist known for his studies of freshwater lakes and considered the father of American limnology....
's famous "Homage to Santa Rosalia" was the next foundational paper in the history of the theory. Its subtitle famously asks, "Why are there so many kinds of animals?", and the address attempts to answer this question by suggesting theoretical bounds to speciation and niche overlap. For the purposes of understanding limiting similarity, the key portion of Hutchinson's address is the end where he presents the observation that a seemingly ubiquitous ratio (1.3:1) defines the upper bound of morphological character similarity between closely related species. While this so-called Hutchinson ratio and the idea of a universal limit have been overturned by later research, the address was still foundational to the theory of limiting similarity.
MacArthur
Robert MacArthur
Robert Helmer MacArthur was an American ecologist who made a major impact on many areas of community and population ecology....
and Levins
Richard Levins
Richard "Dick" Levins is a mathematical ecologist, and political activist. He is best known for his work on evolution in changing environments....
were the first to introduce the term 'limiting similarity' in their 1967 paper. They attempted to lay out a rigorous quantitative basis for the theory using probability theory
Probability theory
Probability theory is the branch of mathematics concerned with analysis of random phenomena. The central objects of probability theory are random variables, stochastic processes, and events: mathematical abstractions of non-deterministic events or measured quantities that may either be single...
and the Lotka-Volterra competition equations. In doing so, they provided the ultimate theoretical framework on which many subsequent studies were based.
Theory
As proposed by Macarthur and Levins in 1967, the theory of limiting similarity is rooted in the Lotka-Volterra competition model. This model describes two or more populations with logistic dynamics, adding in an additional term to account for their biological interactions. Thus for two populations, x1 and x2:where
- α12 represents the effect species 2 has on the population of species 1
- α21 represents the effect species 1 has on the population of species 2
- dy/dt and dx/dt represent the growth of the two populations with time;
- K1 and K2 represent these species’ respective carrying capacities
- r1 and r2 represent these species’ respective growth rates
Macarthur and Levins examine this system applied to three populations, also visualized as resource utilization curves, depicted below. In this model, at some upper limit of competition α, between two species x1 and x3, the survival of a third species x2 between the other two is not possible. This phenomenon is termed limiting similarity. Evolutionary, if two species are more similar than some limit L, a third species will converge towards the nearer of the two competitors. If the two species are less similar than some limit L, a third species will evolve an intermediate phenotype.
[embedded graph: U v R. x1, x2, x3 curves.]
For each resource R, U represents the probability of utilization per unit time by an individual. At some level of overlap between species x1 and x3, the survival of a third species x2 is no longer possible.
May extended this theory when considering species with different carrying capacities, concluding that coexistence was unlikely if the distance between the modes of competing resource utilization curves d was less than the standard deviation of the curves w.
Applied Examples
It is of note that the theory of limiting similarity does not easily generate falsifiable predictions about natural phenomenon. However, many studies have tried to test the theory by making the highly suspect assumption that character displacementCharacter displacement
Character displacement refers to the phenomenon where differences among similar species whose distributions overlap geographically are accentuated in regions where the species co-occur but are minimized or lost where the species’ distributions do not overlap. This pattern results from evolutionary...
can be used as a close proxy for niche incongruence. One recent paleoecological
Paleoecology
Paleoecology uses data from fossils and subfossils to reconstruct the ecosystems of the past. It involves the study of fossil organisms and their associated remains, including their life cycle, living interactions, natural environment, and manner of death and burial to reconstruct the...
study, for example, used fossil
Fossil
Fossils are the preserved remains or traces of animals , plants, and other organisms from the remote past...
proxies of gastropod body size to determine levels of character displacement over 42,500 years during the Quaternary
Quaternary
The Quaternary Period is the most recent of the three periods of the Cenozoic Era in the geologic time scale of the ICS. It follows the Neogene Period, spanning 2.588 ± 0.005 million years ago to the present...
. They found little evidence of character displacement, and they concluded that "limiting similarity, as seen in both ecological character displacement and community-wide character displacement, is a transient ecological phenomenon rather than a long-term evolutionary process". Other theoretical and empirical studies tend to find results that similarly play down the strength and role of limiting similarity in ecology and evolution. For example, Abrams (who is prolific on the subject of limiting similarity) and Rueffler find in 2009 that "there is no absolute limit to similarity; there is always some range of mortality rates of one species allowing coexistence, given a fixed mortality of the other species".
What a lot of studies examining limiting similarity find are the weaknesses in the original theory that are addressed below.
Criticism
The key weakness of the Theory of Limiting similarity is that it is highly system specific and thus difficult to test in practice. In actual environments, one resource axis is inadequate and a specific analysis must be done for each given pair of species. In practice it is necessary to take into account:- individual variations in resource utilization curves within a species and how these should be weighted in calculating a common curve
- whether the resource in question is a present in a deterministic or stochastic distribution and if this changes over time
- effects of intraspecific competition vs interspecific competition
While these complications don't invalidate the concept, they render limiting similarity exceedingly difficult to test in practice and useful for little more than didacticism.
Furthermore, Hubbell and Foster point out that extinction via competition can take an extremely long time and the importance of limiting similarity in extinction may even be superseded by speciation. Also, from a theoretical standpoint, small changes in carrying capacities can allow for nearly completely overlapping resource utilization curves and in practice carrying capacity can be difficult to determine. Many studies that attempt to explore limiting similarity (including Huntley et al. 2007) resort to examining character displacement as a proxy for niche overlap, which is suspect at best. While a useful-if simple-model, limiting similarity is nearly untestable in reality.
See also
- Competitive exclusion principleCompetitive exclusion principleIn ecology, the competitive exclusion principle, sometimes referred to as Gause's law of competitive exclusion or just Gause's law, is a proposition which states that two species competing for the same resources cannot coexist if other ecological factors are constant...
- Competitive Lotka–Volterra equations
- Lotka-Volterra equationLotka-Volterra equationThe Lotka–Volterra equations, also known as the predator–prey equations, are a pair of first-order, non-linear, differential equations frequently used to describe the dynamics of biological systems in which two species interact, one a predator and one its prey...